CO2 proxies based on leaf gas-exchange principles help in part to address the limitations of the stomatal frequency proxies. A well-vetted model (see figure) for carbon assimilation (photosynthesis) describes the rate of carbon assimilation (An) to be equal to the product between the physical capacity of the leaf to exchange gases with the atmosphere (i.e. total leaf conductance to CO2, gc(tot)) and the gradient between atmospheric (ca) and intercellular (ci) CO2 (ca – ci). That is, a higher rate of photosynthesis is associated with a higher leaf conductance and/or a strong drawdown of intercellular CO2 relative to the atmospheric concentration. Rearranging for atmospheric CO2, the equation becomes:

In fossils, total leaf conductance can be estimated from direct measurements of stomatal size and density, along with other parameters that are in part calibrated using present-day relatives. The ci/ca ratio is determined from measurements of fossil leaf δ13C along with an estimate of paleoatmospheric δ13C (for details on this, please see FAQ How exactly do paleo-CO2 proxies work?, in particular the general background information). The assimilation rate (An) cannot be measured directly on fossils and is normally inferred from a nearest living relative, along with a mechanistic model that describes how the assimilation rate scales with atmospheric CO2.
The leaf gas-exchange proxies are largely mechanistic in nature, can be applied to most stomatal-bearing fossil leaves, and produce estimates of CO2 that remain well-bounded (i.e. their upper uncertainties do not go to infinity, as is the case with the stomatal density and stomatal index proxies), even at high CO2.
 Franks leaf gas exchange proxy flow chart
Franks leaf gas exchange proxy flow chart 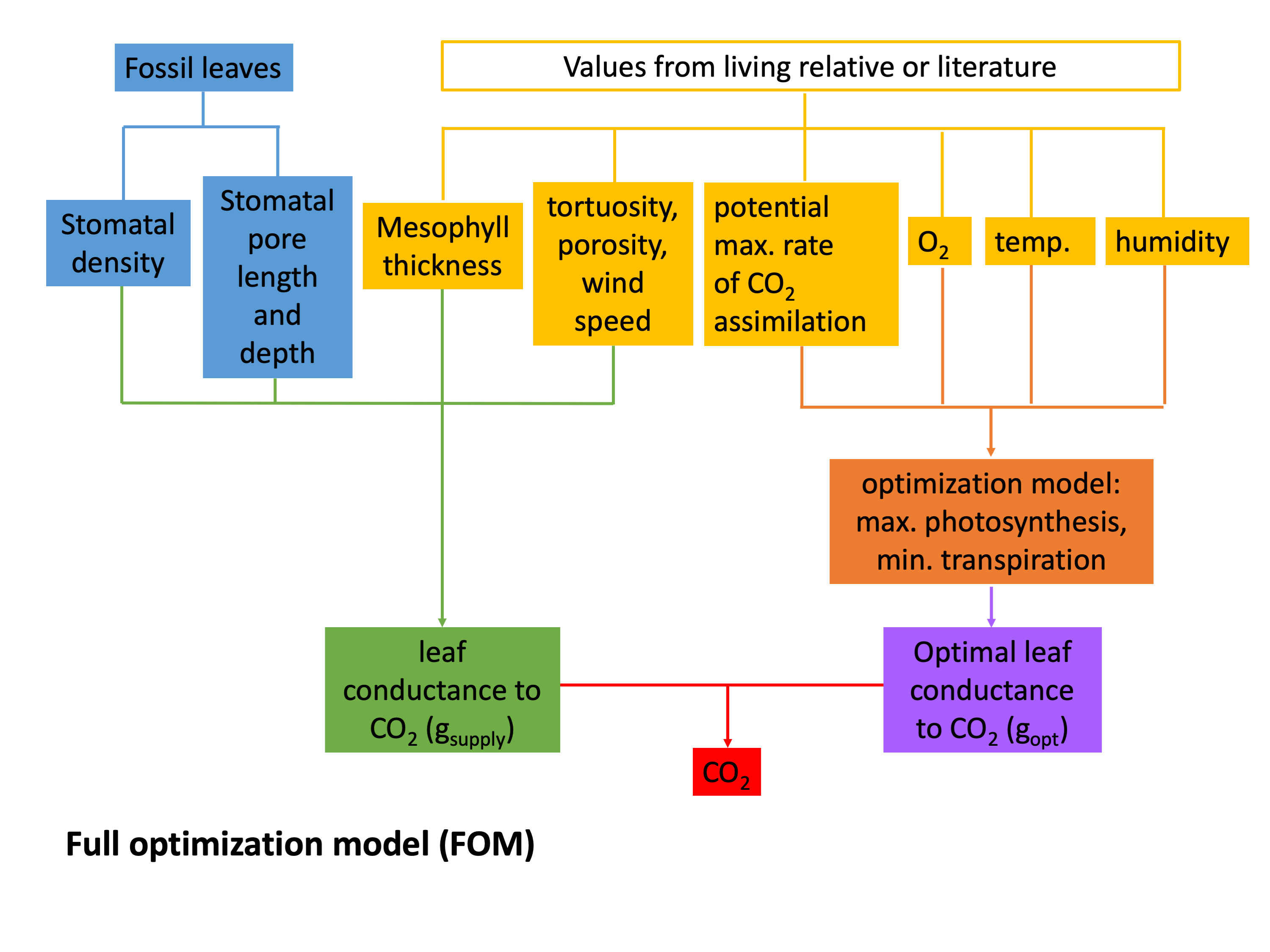 FOM leaf gas exchange proxy flow chart
FOM leaf gas exchange proxy flow chart 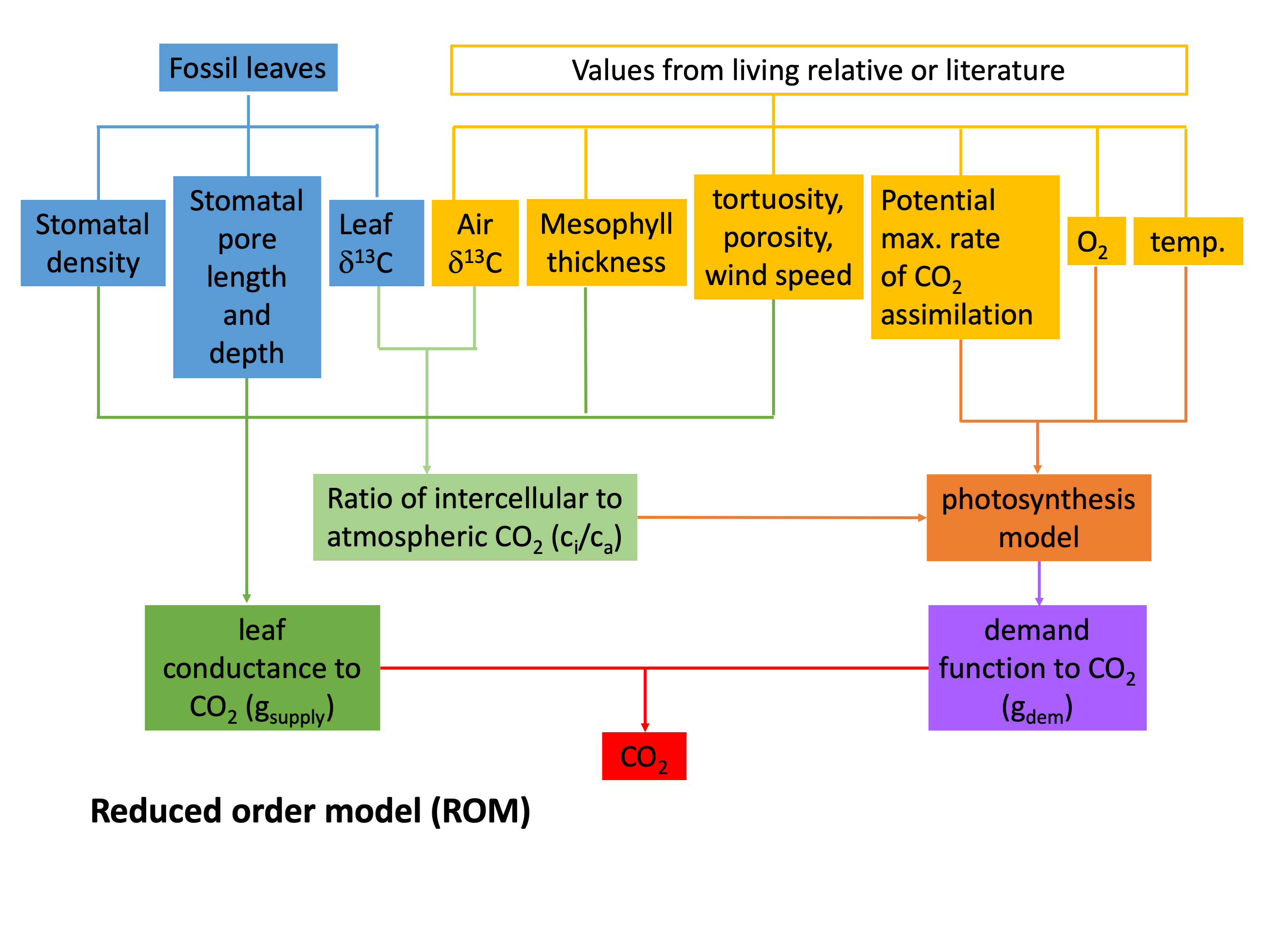 ROM leaf gas exchange proxy flow chart
ROM leaf gas exchange proxy flow chart References cited
Franks, P.J., Royer, D.L., Beerling, D.J., Van de Water, P.K., Cantrill, D.J., Barbour, M.M. and Berry, J.A. (2014) New constraints on atmospheric CO2 concentration for the Phanerozoic. Geophysical Research Letters 41, 4685-4694.